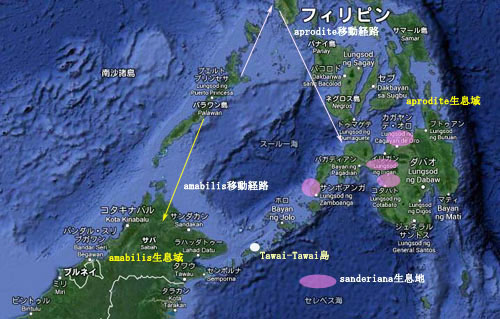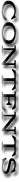
Phalaenopsis sanderiana
|
1.生息分布
| |||||
|
2.生息環境 海抜0-500m、温度20-32C。湿度80-85% | ||
|
3.形状 3-1 花
1. 花被片 花径は5-8.5cm。花被片は全体が白地であり、dorsal sepalおよびpetalの先端部から中心にかけて薄紫のグラデーションが入る。Phal. aphroditeやPhal. amabilisの近縁種である。稀に花被片全体が白色のalbaタイプや、赤紫色あるいは青紫が全体に強く出たもの(左写真)が見られる。生息地の高度が高くなればなるほど花被弁の色が濃くなると言われる。Phal. aphroditeやPhal. philippinensis同様に花茎に2列に並んで多数の花をつける。上写真右の白色の花株は下写真下段の左と中央に見られるように花被片全体が白色で、淡い紫色を帯びたフォームである。本種はPhal. aphroditeとPhal. schillerianaとの自然交配種とされたことがあったが、花被片形状にはほとんど違いがなく、本種には花被片の色が、Phal. aphroditeの白色からP.schillerianaの紫色まで個体差として見られるからであろう。しかし本種の葉にはPhal. schillerianaのような大理石模様はなく、またPhal. aphroditeの無地の鮮緑(青リンゴ)色もほとんど見られない。Phal. aphroditeやPhal. amabilisとの種別はカルス形状で行われる。 最近のDNA分析によるとPhal. amabilisにより近い関係と言われるが、本種が生息するミンダナオ島南部周辺にPhal. amabilisは生息していない(Sulu諸島のTawi-Tawi島にはPhal. amabilisが生息するが、Borneo島に近い)。開花期はPhal. amabilisやPhal. aphroditeが初春に対して夏。しかし温室ではPhal. aphroditeやP.schillerianaよりやや遅い程度で春に開花することも多い。香りはない。花名はF. Sanderから。
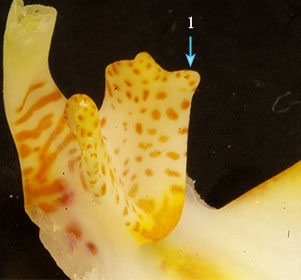
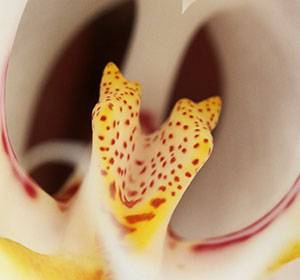
2. リップおよびカルス 本種のリップおよびカルスを示す。本種の特徴はPhal. aphroditeと中央弁の3角形状は似るが、カルスの上部手前(右画像1)が突起状になっていることである。
下画像はPhal. amabilsおよびPhal. aphroditeの中央弁およびカルスの比較を示す。
3-2 さく果 花茎と同じ緑あるいは茶褐色で、花被片は左写真に示すように1ヶ月ほどで縮み枯れる。4カ月程度で播種ができる。
3-3 変種および地域変異 1.Phalaenopsis sanderiana f. alba
2. Phalaenopsis sanderiana dark_purple 赤紫の強いdarkタイプ。フィリピンラン園にて10年の間、数百の本種を見てきた中で最も青紫色の濃いフォームで、発色濃度は再現性がある。
3-4 葉 葉は30 - 35cm x 8cm。下写真に示すように緑色から濃緑色と銀泥色の斑模様などがあり、葉裏は表面と同じ緑色もあるが、多くが中央左画像のような紫色や茶褐色となる。葉は厚みがあり、長楕円形で下垂する。よって小苗時を除き、ポット植えには適さずコルク、ヘゴあるいはバスケット植えとなる。
3-5 花茎 花茎は40cm以上で赤軸が多い。3-6 根 根は太く銀白色。根冠は緑褐色。多数が旺盛に伸長するためコルクやヘゴは大きなサイズが必要となる。 | |||||||||||||||||||||||||||||||||||||||||




















|
4.育成
| ||||||||||||||||||||||
|
5.特記事項 本種は業者のなかでも取扱いが少なく意外と入手しずらい。また注文してもPhal. aphroditeと誤って入荷することがある。葉の色が表裏とも一様な鮮緑色である場合は疑ってみる必要がある。DNA分析によるとPhal. sanderianaはPhal. amabilisに最も近い種とされる。Phal. sanderianaは現在ミンダナオ島西部に生息し、ルソン島をはじめミンダナオ島以北のフィリピン諸島には生息していない。Phal. sanderianaがPhal. amabilisから進化したとする説に従えば、Phal. amabilisはPalawan諸島でPhal. aphroditeから誕生した種であり、この結果、現在のミンダナオ島のPhal. sanderianaは、Palawan諸島からルソン、ネグロス、レイテ島と南下してミンダナオ島に移動したPhal. amabilisを祖先と考えるのが普通である。しかしこの説にはPhal. aphrodite、Phal. amabilisおよびPhal. sanderianaそれぞれのフィリピン諸島における現在の分布状況から考察すると、大きな矛盾が生じる。その最も不可解な要因は、Palawan諸島で進化したPhal. amabilisがミンダナオ島に前記それぞれの島を経由し移動したとすればルソン、ネグロス、レイテ島にもPhal. amabilisが生息している筈である。しかし、いずれの島々にも現在Phal. amabilisは見つかっていない。 Phal. amabilis同様にPalawanから南下した、Phal. aphroditeはフィリピン全域に存在する。Palawanを発祥の地とするPhal. aphroditeがフィリピン諸島全体に分布する一方で、PalawanにてPhal. aphroditeから進化したPhal. amabilisがその経路とされる島々には生息していない。しかしミンダナオ島のPhal. sanderianaはDNA分析によればPhal. aphroditeからではなくPhal. amabilisから進化したのである。よってミンダナオ島でPhal. sanderianaを誕生させたPhal. amabilisはどこから来てどこに消えたかという疑問が残るのである。 ルソン島からミンダナオ島に分布するPhal. fasciataやlueddemannianaなど多くのフィリピン固有種は5百万年前のフィリピン諸島がボルネオから分離した後に誕生した種であるためフィリピン諸島内の固有種として全体に広く分布生息している。これらがフィリピン諸島に分布した過程はルソン島からミンダナオ島に至る南下説が有力となっている。一方、フィリピンはルソン島とボルネオ島を結ぶPalawan諸島とは別に、もう一つミンダナオ島からボルネオ島に繋がるSulu-archipelagoにも列島(下写真参照)があり、その列島の中でボルネオ島寄りとなるフィリピン領の小さなTawai-Tawai島にはP. amabilisが現在生息していることが知られている。すなわちフィリピン領内でのPhal. amabilisはいずれもボルネオ島を結ぶPalawanとTawai-Tawai島の2か所にのみに生息している。 Phal. amabilisはPalawanからボルネオ島に移動し、さらにSumatra島やJava島へ移動していることから、Palawanからルソン島経由の南下説ではなく、ボルネオ島からSulu-archipelagoを迂回してミンダナオ西部に至ったのではとする北上説が考えられる。このPhal. amabilisの移動説は現在の学説にはない。 ボルネオ島にはPhal. aphroditeは生息せず、またミンダナオ島にはPhal. amabilisは生息していないことを考えればボルネオ島に近いTawai-Tawai島のPhal. amabilisはボルネオ島から移動したものである可能性が高くなる。すなわちPhal. amabilisはPalawanからボルネオ島へ、さらにSulu-archipelagoを北上し、ミンダナオ島周辺(Zamboanga)に達して、Phal. sanderianaが誕生したのではと推測される。この時期同時に起こった氷河期が終わり海面上昇により、フィリピン諸島がPalawanやSulu-archipelagoを挟んでボルネオ島から分離され、この結果、Phal. amabilisはボルネオ島寄りのSulu-archipelago西端に、またPhal. sanderianaはミンダナオ寄りの東端に残ったとすればルソン島やミンダナオ島にはPhal. aphroditeが生息していてもPhal. amabilisが生息していない理由が説明できる。Phal.aphroditeとPhal. amabilisの内、Phal. aphroditeだけがフィリピン諸島に広く生息するのは、Phal. amabilis誕生以前の陸続き時代に分布したものと考えることができる。
現在のスラウエシ、パプア・ニューギニア、オーストラリアに生息するPhal. amabilisはPalawan-ボルネオ島-スマトラ島-Javaルートではなく、Palawan-Luzon-Mindanao島を経て移動してきたとされている現在の学説も同様の疑問点があり、Phal. amabilisがフィリピン諸島を南下してそれらの地域に移動したのであれば、ルソン島、ミンダナオ島をはじめとするそれぞれの島にPhal. amabilisが生息していると考えられる。しかしこの実態がない以上、Phal. amabilisのフィリピン以外の地域への広域分布の起点は全てボルネオ島であり、ボルネオ島からスラエシ島、マルク諸島あるいはJava、ティモール、マルク諸島を経て分布した地理的には西に迂回したルートの方が論理性がある。これらスラエシ、Java、マルク諸島には現在それぞれPhal. amabilisが生息している。 上記の考察は2つの学説;Phal. amabilisはパラワンにてPhal. aphroditeから進化したとする説と、Phal. sanderianaはPhal. aphroditeよりもPhal. amabilisに遺伝的距離が近いとする説を前提としている。しかしPhal. amabilisがパラワンにて誕生し、ボルネオ島に移動したことを是としても、Phal. amabilisではなく、Phal. aphroditeがルソン島やレイテ島を経てミンダナオ島へと南下し、ミンダナオ島にてPhal. aphroditeからPhal. sanderianaが誕生したとする仮説はどうであろうか。興味のあることはPhal. aphroditeのカルスとPhal. sanderianaのカルス形状は前記3-1項の’リップおよびカルス’での画像から分かるように酷似している一方、Phal. amabilisのカルス形状とはむしろ離れており、遺伝的距離が近いとされながらもカルス形状の差はその逆を示している。すなわちカルス形状からみればPhal. sanderianaはPhal. aphroditeから進化したように思われることである。さらに面白いことは現在Phal. sanderiana f. albaとされる種の存在である。アルバ変種の出現は、数万株に一つとされながらも、不思議なことに野生株でありながら、ここ10年ほどの間にミンダナオ島野生種として相当数を、信頼あるサプライヤーから入手している。なぜこれほどの野生アルバ株が入手可能なのか。一つの仮定はこのf. albaはPhal. sanderianaのalbaフォームではなく、Phal. aphroditeから進化した別種でミンダナオ島に広く分布する固有種であり、この種がPhal. sanderianaの誕生に関っているのではないかとも考える。これにはf. albaのミンダナオ島内陸における分布実態を調べることが必要で、その解明(Phal. amabilis北上説を含め)にも重要な地域であるミンダナオ島西域は、現在治安の問題で立ち入ることが出来ないが、それが可能となれば様々なことが解き明かされるのではないかと思われる。 生息分布の地理学的考察と、種の特徴を示すとされるカルスの形態学的視点からは、このような様々な疑問や仮説が生まれる。しかし、こうした推理を巡らすこともラン栽培者としての楽しみの一つである。 | |||