|
花形状 胡蝶蘭の花は、花被片、唇弁(リップ)および蕊(ズイ)柱の3つの部位に分かれ、花被片は写真1に示すように背ガク片、花弁および側ガク片から、さらにリップは中央弁、側弁およびカルスを、また蕊柱は花粉魂のそれぞれからなります。花被片の形状は多くの種で星形であり、写真左のような園芸店でよく見かける白色の大きな花弁をもつ胡蝶蘭は全体から見れば10%程度の少数種となります。花の各部位は写真1に示す名称となっています。50種以上ある胡蝶蘭原種の花柄は多様であり、また他属と同様に多くの種でalba(白化), flava(黄化)が, また一部にcoerulea(青色)の変種が見られます。自然界において、これらの変種は数千あるいは数万に一つの確率で存在し、実際に自然界において見ることは非常に困難とされ、今日市場にあるのは、過去発見された株を元に、それらのメリクロンあるいは実生が大半です。 花の同時開花数は、2-3輪を半年間に渡って次々と咲き続ける種(P.cornu-cervi等)から、数10輪をほぼ同時に開花させる種(P. amabilis, P.schilleriana等)まで多様です。 一方、花期は同じ温室内の栽培でありながら年1回、春秋の2回また不定期など、それぞれがあり早春から初夏に比較的多いものの、属全体としての一つの定まった開花期はありません。栽培では生息地と温室の環境の違いで自然とは異なる開花期も見られます。 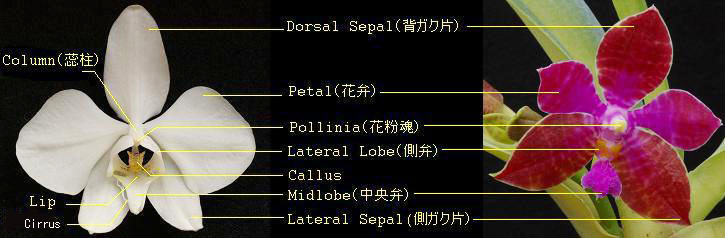 Fig. 1. Phalaenopsis Flower (Left: P.amabilis, Right: P. lueddemanniana f. mindanao) 写真2にはリップを中心に拡大した数種類の花形状を示します。リップは花柄と同様に様々な形状をもち、いずれも左右対象の側弁(lateral lobes)と、中央弁(midlobe)および中央弁の基部にあるカルス(callus)で構成され、リップ先端部の小裂片(lobules)は下記の6つのタイプに分類できます。
 Fig. 2. 胡蝶蘭リップ形状 | ||
|
開花時期 表1には原種の開花時期を示します。複数の季節にまたがるものや、国内の温室栽培では開花時期が自然界とは大きく異なる場合もあります。
開花期間は数か月(1-2ヶ月)に及ぶもの、花単体の寿命は2-3週間でありながら数か月(<em><a href="../../species/cornu-cervi/specieshead.html"target="_top">P. cornu-cervi</a></em>)あるいは半年以上に渡って次々と開花を繰り返す(<i><a href="../../species/equestris/specieshead.html"target="_top">P.equestris</a></i>の一部など)ものがあります。栽培環境においては、移植年度、水分欠乏、根ぐされ、栄養不足等による弱体化が生じている場合は、4-5日で萎れることがあり、また葉が2枚程度しかない株や、実生からの初花では、花柄は小さく輪花数も少なくなります。 | ||||||||||||||||||||
|
リップとカルス カルス(callus)はリップの基部(付け根)あるいは基部から突き出た突起状のものをいい、写真左のP. amabilisように一つの塊からなるもの、中央の先端が2つに分岐した歯状突起(anteriorおよびposterior)をもつもの、さらに 右端写真のように髭状(anterior)のものから小さな凹凸のある腺状突起(posterior)をもつものがあります。写真はいずれも、カルスを見やすくするためリップの側弁を切り取っています。カルスの多くはanterior(前方)とposterior(後方)の2組で構成されます。anteriorは殆どの種で先端を2分岐した歯状突起をもち、posteriorは2分岐歯状か腺状突起となります。これらのカルスはそれぞれが独立したり連続するものなど種で多様に変化し、花被片(花びら)の形状、色、模様などからは種の同定が困難な場合の重要な判断要素となります。 Fig.3. 各種カルス (左からamabilis, modesta, lamelligera)。側弁を切り取った状態 | ||
|
蕊柱・花粉魂・さく果 写真4に蕊柱の拡大写真を示します。(1)は先端部を下方から見た映像です。中心部には大きな窪み(雌しべに相当)があり、先端部には花粉魂を保護する葯帽(シャクボウ:anther cap)と、葯帽の内側からはリボン状粘着体(rostellum)が窪みを横切るように飛び出しています。写真(1)では葯帽の裏にうっすらと黄色の花粉魂(pollina)が透けて見えます。リボン状粘着体(学術的には上段左端のリボン状粘着体を指す部分を小嘴(シ)体といい、上段右端のリボン状粘着体を指す部分を花粉魂柄(stipe)という)の一方は花粉魂と接着しています。リボン状粘着体(小シ体)の内側には強力な粘着物質があり、引っ張られると粘着物質がゴムのように伸びてその長さは粘着体の3-4倍になります。昆虫がこの飛び出たリボン状粘着体に触れると瞬時に接着し、動けば花粉魂と葯帽が蕊柱からポロリと外れます。写真(2)は葯帽、花粉魂、粘着体が外れた状態を示します。リボン状粘着体の外れた場所には爪状突起(talon)が残っており、これが受粉の際に花粉魂を窪みに留める重要な役割を果たします。写真(3)は葯帽と花粉魂をそれぞれ示したものです。花粉魂と葯帽自体は粘着性はなく、葯帽は花粉魂にまさに帽子のようにすっぽりと被っている状態で、昆虫が移動したりして振動を与えれば葯帽は花粉魂から離れ落ち、黄色い花粉魂とリボン状粘着体(4)だけが昆虫に残ります。昆虫が再び蕊柱の窪みに近づき、また出ようとするとき花粉魂は窪みの内壁(雌しべ)にはまります(5)。花粉魂とリボン状粘着体は、写真(4)のように1対の花粉魂の一角をセロテープでとめたような状態で付いており、引っ張れば剥がれる構造となっています。この花粉魂をチョンと人工的に仮着けしたような構造への進化には自然の驚異を感じさせます。 窪みの壁自体には花粉魂を付着する程の粘着物質はなく、リボン状粘着体と花粉魂との接着力の方が遙かに強力で、窪みに花粉魂が収まったとしても昆虫から花粉魂が離れる力はありません。ここで写真(2)の爪状突起が鈎となって、花粉魂が窪みに入ると、昆虫が離れる際に花粉魂を引っかけると共に、爪状突起の根元に引き寄せます。この根元には粘着物質があり花粉魂をしっかりと窪みの中(突起の裏側)に抱き込む形となります。昆虫が力づくで離れようとすると、粘着体はゴムのように伸びて昆虫と、爪状突起を挟んでの引っ張り合いとなり、粘着体が昆虫から離れるか、粘着体は昆虫に着いたまま花粉魂のみが窪みに残るかのいずれかになります。写真(5)には窪みに花粉魂とリボン状粘着体が共に残った状態(受粉完了)を示します。自然界の受粉では、昆虫側に付いたリボン状粘着体の接着力は極めて強く、花粉魂だけが窪みに残る確率が多いのではないかと推測されます。しかしリボン状粘着体が残っても受粉自体には影響がないようです。2‐3日で窪み部分は写真(6)に示すように花粉魂を巻き込んだまま閉じられていきます。この状態から凡そ4ヶ月で種子(胚生成)が完了します。右はP. violaceaの花粉魂を拡大した写真で、粘着体と花粉魂が糊状の物質で接着している部分が見えます。  Fig. 4 蕊柱先端部と花粉魂
特にP. lindeniiなど一部の種では、高温下におくと受粉から1週間ほどで萎れて落下してしまうことが多く、また種によっては花粉からのホルモン不足が原因と思われる現象が見られます。このような場合はホルモン剤オーキシンを花粉魂とともに蕊柱に滴下します。 一方、受粉によってやがて子房は膨らみ長く延びます。この受粉によってできる果実部分を本サイトでは「さく果」と呼ぶことにします。交配後、花被片(花びら)は縮れて枯れる種(phalaenopsis亜属)と、さく果と共に緑色に変色して硬くなり、さく果が割れるまでそのままの状態となる種(polychilos亜属)があります。またさく果の外皮の色も緑色、茶褐色(stauroglottis節)、その中間(phalaenopsis亜属の一部)などが見られます。さく果は下写真のような縦に6つの筋状の凹みをもちます。前記したように交配から4か月程で胚が完成し、種子の取り撒きが可能となりますが、そのまま放置しておくと、環境条件によりますが6か月から10か月程度で黄変し、黄変し始めると4-5日で子房の一部が割れます。割れる部分は種が容易に落下するように、常に子房の下部で起こります。どうして上下の判断ができるか面白い特性です。下図のそれぞれに一部の種のさく果を示します。写真映像のサイズはスケール対比をしていません。図中ではP. giganteaがもっとも太く大きなさく果となります。  Fig. 6 さく果形状 | ||||

|
虫媒花 同じ属でありながら、写真2に見られるようにリップ形状、色およびテキスチャ‐がそれぞれ異るのは、受粉に必要なそれぞれの種に固有のポリネーター(受粉媒介者)を招くためと、停まりやすい形に進化したためと考えられています。また僅かな輪花数でありながら長期間次々と咲き続ける種や、短期間に多くの花を一斉に咲かせる種があり、この特性の違いは、いずれもポリネーターの発生特性やその数に対して受粉確率を高める共生関係を得るための進化と思われます。花粉魂はリボン状粘着体に触れるもの何ににでも瞬時に接着します。この何にでも取りつく性質は反面、個体数の少ない蘭にとって受粉確率を低下させることになり、種の存続を危うくすることになります。また、もし胡蝶蘭の受粉が「特定の」虫媒介でなければ、これほどの複雑な構造に進化する必然性も見当たりません。これらから同じ種(あるいは属)をターゲットに飛び回る限られたポリネーターがいることによって、一定の受粉確率を高めていると推定することができます。さらに花粉構造の条件から花粉媒介者は蜂あるいはハエのような羽を持ち、また蕊柱やリップ側弁を潜り抜けることのできる大きさの昆虫と考えられます。蝶(花粉魂接着場所と受粉の問題)や羽の無い虫(虫の移動距離と蘭の個体数の問題)による受粉の可能性は少ないと思われます。 一方、ポリネーターは何に誘われて胡蝶蘭の蕊柱に飛来するのでしょうか。カトレアやラリア(例えばアンセプス)は匂いや蜜線からの粘着液が主な誘引物質の一つと考えられますが、胡蝶蘭ではポリネーターの餌となる花粉も、蜜のようなエネルギー物質もなく、また多くの種が微香であり、全体として見ると香りの有無はまちまちです。それ以外の要因としては花の形が擬態という見方もあります。Parishianae亜属のリップの形状、色あるいはテキスチャ‐を見ていると、擬態らしい推測もできます。しかし全種がそうであるかと言えばこれも疑問です。これらの様態を考えれば、属としてポリネーターを誘引する共通した物質(餌、色、擬態などの一つ)を確定することが困難です。 花粉を虫が運ぶ進化は一説によると、まずパウダー状の花粉を付けた植物が最初に現れ、風で花粉を飛ばして受粉を行っていましたが、少数小型の種にとって、この風任せの方法は、極めて受粉効率の悪いシステムであり、ほとんどの花粉が無駄になります。やがて昆虫のなかに花粉を食べるものが現れ、植物にとって花粉が食べられてしまうのは損失であるものの、虫は確実に花の間を行き来することから、虫の体に付着した花粉が仲間の花に運搬される確率は風よりは遙かに高く、この結果、子孫をより多く残すことができ、風媒花からこのような虫媒介のシステムが出来上がったとされます。 そうするとその植物の中には、花粉を食べられることなく虫に運んでもらおうと進化した植物が現れ、風で飛ぶことも食べられることもない固い花粉魂を葯帽と粘着体で包み、一方、適度な蜜(多いと昆虫は一花で満足し、他の花へ行かない)や匂いを出して虫を呼び寄せ、虫の体に花粉魂を接着させて一層確実に花粉を運ばせるようにします。さらに進化した植物では、植物にとっては蜜や匂いを出すことが相当な体力を必要とすることから、蜜も匂いも出さず、形や色で昆虫を欺き、あるいは特殊な成分を雌しべに僅かに分泌して虫を呼ぶものまで現れるに到ったという訳です。 胡蝶蘭に関してはポリネータ‐の研究が見当たりません。アツモリソウ(シペリペヂウム)はそれぞれ種によって異なる匂いを出し、これとリップ形状のサイズから特定のポリネータ‐(マルハナバチ、ミツバチ、ハナバチ、アブ、ハエなど)を選択的に誘引するようです。しかしパフィオペディラムはこの差別化した匂い成分がなく、現状では色や形ではないかとされているものの、確定的な誘引物質が良く分かっていません。中南米での一部の蘭は、蜜を出さない、擬態もないにもかかわらずハチ(シタバチ)のオスを誘引するそうです。オスは花の中(胡蝶蘭では腺状突起カルス周辺あるいは蕊柱の窪みに相当)にある物質(香料の原料となるシオネール成分などを含む)を前足でかき集め、後足のスポンジ状の袋に蓄えるそうです。この芳香物質をオスだけが求める理由は、唾液と混ぜ合わせてメスを誘引するフェロモンを生成するためと言われています。つまりこの蘭は花粉や蜜を提供するのでも、密やメスを真似た香りを出す訳でもなく、蜂のメスを呼び寄せるフェロモンの原料をオスに提供している訳です。恐るべき共生の進化と言えます。 最も進化したCAM植物である胡蝶蘭は研究報告がない以上推測でしかありませんが、種に応じてそれぞれ誘引手段の主体が変化し、主にamboinenses亜属は匂いが、parishianae亜属は擬態を、またその他の亜属は色、テキスチャ‐あるいは前記それぞれの要因のAnd/Orが引き金ではないかと思われます。餌や蜜が胡蝶蘭には無い以上、繁殖目的のフェロモンの生成物質を提供しているのかも知れません。 | ||
|
香り 胡蝶蘭およそ50種の内、他の蘭属と比較して香りを持つものは少ないと言われています。香りをもつ代表的な種はP.violacea、 P.lueddemanniana、 P.modesta、 P. floresensis, P. sumatranaなどが知られていますが、開花後からの一定期間あるいは時間帯によって、意外と匂いが無いとされている種でも微香があります(P.amabilisなどの一部地域のものは青草のような香り)。種によっては匂いも、種や地域差を同定するための重要な要素となります。例えば今日、P.violaceaやP.bellinaの純正な野生種(山採り)を得ることは非常に難しく、同種間や異種間で、ソリッドな青や赤系色のP.violaceaを得るために、選別交配が進み自然界では起こりえないような色彩を持つものが多く出回っています。野生種か交雑種かの1つの判断として、P.violaceaやP.bellinaにおいては、野生種のP.bellinaは強いレモンの香りをもっており、一方のP.violaceaではスパイス系の香りがあり、同時に比べて見ると違いが良く分かります。P.bellinaとP.violaceaとの交配で花弁の形はP.bellinaと同じであっても、レモンの匂いがなければ、これらの交配種と見なしてもほぼ間違いがないものと思われます。香りで不思議なことは、例えばP.modestaのように同一種でありながら地域によって匂いのある(ボルネオ島)ものと、匂いを感じない(スマトラ半島)ものがあることです。これはP. cornu-cervi系にも見られます。香りの違いはそれぞれ異なるポリネータ‐を誘引ためであることを考えると、一方の地域に生息するポリネータ‐が、一方には生息しないため、それぞれの環境のポリネータに合わせて香り成分が進化したのか興味のあるところです。多くの胡蝶蘭は明るい時間帯に香りを出し、夜は香りを止める性質があります。特に午前中が最も香りが強くなります(P. kunstleriは夜に香りを放ちます)。
人にとって好ましい香りかどうかは個人差によるため香りの評価は一概にできませんが、個人的な嗜好からすれば良い匂いと感じるのはP. violacea、P. lamelligera、P. hieroglyphicaおよびP. giganteaでしょうか、柑橘系の香りがします。これに対して P. corningiana, P. floresensis, P. sumatranaは漆喰(昔の粉石鹸と言う人もいます)匂いのようで好きではありません。 | ||||||||||
|
葉形態 熱帯モンスーン気候に生息する着生植物にとって乾季の乾燥を凌ぐためには、葉、茎および根のいずれかに水分を蓄える必要があります。カトレアやラリア、また同じ東南アジアに広く分布するBulbophyllum等の多くは、主に茎(Bulb)に水分を貯蔵しますが、茎に貯水力のない胡蝶蘭は、葉あるいは根がその役割を担います。このため多くの種の葉は多汁性で厚みがあり、表皮は水分の蒸散を防ぐロウ質(クチクラ層)となっています。葉は水分蒸発の最も多い部分であるため一部には、乾季あるいは寒冷期の間は落葉する種が含まれます。胡蝶蘭は、その生息する環境によってこれら2つの形態に別れます。季節林や、標高が高く寒冷期のある地域では、根が発達する反面、2-4枚ほどの小さな葉となります。 P.wilsonii, P.honghenensis, P.braceanaなどの落葉種がこれらに含まれます。興味のあることは、温室などで定期的な潅水をしていると、これらの種に落葉しない(半落葉)株が現れ、人工栽培では10C以下という環境や、乾季などの季節感が生まれないため水分ストレスが生じないことが原因と思われます。葉は水分を、根は光合成で生成した炭水化物を貯蔵する多くの胡蝶蘭は、根が相当にダメージを受けて(採取の際の根切りや、搬送の際の乾燥)も、葉の状態が良ければ、作落ちはあるものの植え付け後の管理(湿度の保持)で問題なく再生します。それに対して葉が相当なダメージを受けている場合は、落葉種を除いて、発病の可能性を含め再生が極めて困難です。一方、P.modestaやP.fimbriataなどは胡蝶蘭のなかで最も薄葉であり、また落葉種でもありません。通年、湿潤な熱帯低地林に生息しているための進化と考えられます。ほとんどの種で葉は、新鮮な緑色で、卵、楕円および長楕円形をなし、葉裏がやや表面よりも黄味を帯びているものが多く、一部にはP. sanderianaやP. philippinensisなどの茶褐色、また銀色と緑色との斑模様をもつ種(P. schilleriana, P. celebensisなど)があり、これらは葉そのものが鑑賞の対象にもなります。 写真7にはそれぞれの代表的な葉の形態を示します。左上はP.thalebanni、左下がP.bellina f. coerulea、中央がP.schilleriana、右がP.gigantea (kalimantan)です。P.giganteaは葉長が50cm以上になります。胡蝶蘭は単茎性の着生蘭であり、茎は真上あるいは水平それぞれの成長方向があり、それに伴って葉の伸長方向も異なってきます。 P.thalebanniやP.cornu-cerviの茎先端は直すぐ上を向く立ち性ですが、多くの種では、コルクやヘゴ板などの支持体の垂直面へ取り付けられた場合、葉は下垂あるいは水平方向に伸びようとします。 茎が真っすぐ上に伸びる場合、葉は左右交互に開いて展開し、葉の表面は真上を向きます(P. cornu-ceriviなど)。この場合、下葉のほとんどは上葉の真下となりますが、左右交互であるため、それぞれ上下の葉にはある程度の隙間ができ、斜光であれば下葉にも光が届くようになります。一方、水平に伸びる茎では、葉は垂れ下がり、古葉と若葉は前記P.cornu-cerviなどとは異なって、それぞれが密着します。しかしそれぞれの葉は左右に角度を変えて下降するため、こちらでも写真にも見られるように古葉も新葉同様に光を浴びることができます。  Fig. 7 Leaf Morphology 太陽光が真上から降り注ぐ時点では、垂直茎の最上部の葉は太陽光をほぼ直交して受けることになります。一方、下垂する葉では太陽が真上にあるとき光は葉表面と平行となり直射を避けられます。また太陽が斜めから射す場合には他の周辺の林に遮られます。このことから推測されるように、P.cornu-cerviは熱帯季節林や低地雨林に主に生息し、林を抜けた太陽光を出来るだけ多く受け取るための様態となり、一方、P.giganteaなどの下垂する種は林冠あるいは強い太陽光を受けやすい場所に生息し、最も照度の高い昼間の光線は避けるように進化したのではないかと考えられます。
表3には葉の厚み、表4には下垂する葉とアンジュレーション(波打つ)種を示します。種によっては地域差で葉の厚みが異なる場合があります。例えばP.equestrisではroseaタイプは小型で薄葉ですが、leucaspisタイプは全体に大きくP. cornu-cervi並みに厚葉です。
胡蝶蘭の葉の模様は大別して無地と大理石模様の2種類に分かれます。さらに無地は緑色と緑茶褐色となります。大理石模様および無地の緑褐色は表4となります。写真9は左からそれぞれcelebensis, lindenii, philippinensis, schillerianaおよびstuartianaの大理石模様を示します。これら大理石模様をもつ種はすべて下垂タイプの葉をもちます。
 Fig. 9 大理石模様葉種
葉の厚みは栽培上の重要な情報を暗示していると考えられます。薄葉の多くは中光で、湿度が高い環境(あるいは乾燥地帯では落葉する)で生息しているものと思われます。逆に厚葉の種は乾燥に強く、下垂することで太陽光の直射を避けていることになります。これは栽培上、照明が低くても良いということではありません。栽培において下垂状態で且つ、光量を下げれば一層輝度が低下します。例えば、林冠では太陽光を浴びて明るく、また湿度は低く、一方、昼夜の温度差をもち通風が十分にある環境となります。この結果、葉は水分を貯蔵するために厚くなり、また太陽光の直射を避けるため下垂するという進化を経たと考えられ、このような種には十分な照明と比較的強い通風、夜の高湿度化などが理想的な環境であると推測できます。 | |||||||||||||||||||||||||||||||||||||

|
根 胡蝶蘭の根は多くの面積で空気に接しながら水分や栄養分を吸収する「気根」であり、根を取り巻く環境(栽培ではミズゴケ、コルクあるいはミックスコンポストなど)で3つの様態があります。(1)支持体がなく空中に伸びる根は円柱形で表皮は艶のある白色、先端の5-10mm程の根冠だけが新鮮な緑色あるいは緑褐色の色をもち伸長してしていくもの。(2)コルクやへご板などに見られる支持体に接する部分は平たく、空気に触れている部分が皺のある蒲鉾型となるもの。(3)空気に触れている部分が扁平(リボン状)になるもののそれぞれです。(2)はP.cornu-cervi, P.pantherianaなど太い根をもつもので、(3)はP.schilleriana, P.stuartianaを始めとする多くの種がこれに属します。根が活着でリボン状になる種では葉が下垂するタイプが多く、コンポストはコルクやヘゴ板(棒)が適します。一方、太く円筒形あるいは活着後に蒲鉾形となる種はポット植えにも対応できるものが多い印象を受けます。 支持体への活着は根の先端部(根冠およびその周辺)に1-2cm程の帯状に群生する0.5mmもない短い繊毛突起物がコンポストに食い込む形で取り付きます。この突起物は照明が明るい表皮の反対側(裏側)に発生し、支持体がない状態では根が伸長すると共に無くなって、先端周辺だけに新たな突起物が生成されていきます。 ミックスコンポストの植え付けでは、植え込み材の密度によっては地中相当の疑似環境となりますが、P.schillerianaのように、支持体があればリボン状の形状をもつものでも、ミックスコンポスト内では根の表皮は半透明な乳白色で円柱状になります。この根をコルクに植え換えると、主茎からの新根だけでなく、根の先端あるいは根の途中から新たに分岐して生まれた側根だけが支持体に活着し、扁平に変化します。支持体から離れてしまって一度成長した根の部分は再び活着することはできず、活着できるのは根冠部分が支持体に接して新たな細胞が作られる部分に限られます。
胡蝶蘭の根は湿度に屈正を示すとされています。そうであれば、ヘゴ板のように板の下部の湿度が上部より高いものでは、根は下方に向かうと思われますが、筆者の経験からは、そのような傾向を示したのはP.modestaのみで(一定量の上部の根が活着した後、下方に向かう根が増した)、特に葉が垂れる種は、まず上部に向かう傾向が観察されます。垂直に近い面で、下方に根を伸ばしていては自身の体重を支えることはできないため、上部へ延びる傾向が優先するのではと筆者は解釈をしています。また根の屈光性(明るい方向に伸びる)はないとされています。
| ||||||


|
花茎(花序) 胡蝶蘭の花茎は、根と同様に葉の基部(主茎)を突き抜けて現れ、節とえき芽を形成しながら細長く成長していきます。屈光性があり明るい方向に伸長します。特に屈光性の傾向はPhalaenopsis属Phalaenopsis節に顕著な印象を受けます。花茎下部の節にある幾つかのえき芽は休眠し、上部のえき芽が花芽になります。また一部のえき芽は幼苗(高芽)となることがあります。この高芽の出る確率は山採り株(野生種)に高く、特にP.pulchra, P.pallen, P.equestrisなどでは頻繁に見られます。形状は、ほぼ等しい間隔の節をもち上部に長く伸びるもの(P.amabilisなど), 多数分岐するもの(P.schillerianaなど)、弓状に下垂するもの(P.equestrisなど)、円柱形(P.sumatranaなど)や扁平形(P.cornu-cerviなど)があります。花茎発生環境条件の研究はPhalaenopsis亜属のPhalaenopsis節(P. amabilisやP. schillerianaなど)を原種として改良された園芸交配種では国内の研究者らにより詳細に解明されています。これら研究によると25C以下12時間以上の照射によって6-8週間で花茎が発生することが報告されています。特に興味があることは花茎の伸長には昼間の低温が必要で光量は多い方がよく、光照射が長いと花茎数が増加し、夜間の温度が18Cより高く(21C以上)なると花茎数は減少するとされている点です。また昼間28Cでは夜間18Cであっても花茎発生は抑制され、昼間30C、夜間21Cとなると発生が見られないとのことです。 野生種の場合、Phalaenopsis節だけでなく、多くの種がそれぞれ多様な環境に生息しているため上記の条件がどの範囲まで適用できるかは不明ですが、2月から3月にかけて開花するP. amabilisグループでは上記の結果に符合しています。やや遅れて4-5月に開花する種、盛夏に最盛期を迎える種など様々であり、前記の昼間と夜間の絶対温度値が全てに共通となるわけではありません。P. wilsoniiなどのaphyllae亜属では10Cほどの低温に1ヶ月以上置かなければ花茎の発生は抑えられますし、多くの種(例えばP. bellina)が花茎を発生する環境は昼間28C以上夜間20C以上となっていることが観測されます。しかしいずれも共通していることは光照射量と、昼夜の温度差がある程度必要であることです。 原種には交配種とは異なり、しばしば高芽が発生します。花芽となるべき芽が栄養芽に変わるのは、期待していた温度が高くなってしまったためと考えられます。低気温で花茎を出したものの途中で気候が変化し昼間の温度が上昇したり、日照時間が減るなどによって高芽になることがあるそうです。原種の中でも、これらの高芽発生傾向は海外から入荷した株の初年度から次年度に多く、おそらく国内外との環境の変化によって、順化の過程で、これらの種が高芽となる温度状況下に置かれた可能性があります。 P. schillerianaを見ていると、遅れて発生した花茎は開花期を超え高気温にさらされます。この状況となると高芽を付けます。この高芽の毎年の発生はやがて株を大株にします。花は花粉媒介者(昆虫)の発生に合わせて開花している筈ですから種の繁栄にとっては開花時期が最も重要です。不幸にして季節外れとなれば、花がダメなら高芽でもって繁殖するという蘭のしたたかな生存力、自然の驚異を感じます。
花茎の長さも種によって多様です。写真13は4種類の花茎を示します。左上はP.micholiziiで花茎は胡蝶蘭の中では最短で、葉の基部から僅か1cm程度伸び出しています。写真は受粉の結果、花房が成長している状態です。また左下はP.lamelligeraです。扁平の20cm程の花茎に左右交互ジグザグに花を着けていきます。また中央はP.violacea v. mentawaiで50cm程の長い花茎をもちます。胡蝶蘭の中で最も長い花茎は右写真のP.pulchraで、1mを超えます。 花茎も種判定に使用されることがあります。Mentawai島に生息するP.violacea v. mentawaiの花は淡い赤紫色をしていますが、P.violaceaは現在、地域間の交雑が進み、純正なmentawaiを入手することが困難になりつつあるそうです。mentawaiは他のP.violaceaとは異なり、花茎を50cm以上伸ばすことから特定できます。同様にP.pulchraは1m以上に花茎を伸ばす特質があり、もしこれらの種名でありながら短い花茎(初花となる若い苗は除く)であれば、交雑種かと思われます。交雑化が進み、花の形態からだけでは純正種かどうかの判断が近年益々困難になっています。  Fig. 13 花茎の長さ | ||||

|
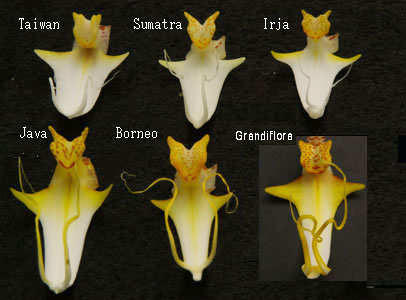
花形態の地域差 胡蝶蘭の中には、生息領域が極めて狭い種(P. inscriptiosinensis, P. javanicaなど)と、広く分布する種類(P. amabilis, P.cornu-cervi、P.sumatranaなど)があり、広域分布種では生息地域特有の花形態が、また地域間境界では自然交配と思われる複合した特徴をもつ株が見られます。しかし、サイズや輪花数については同一固体であっても年毎の環境や施肥条件によって変化します。また個体差を考慮すると花柄や花形状だけで種を判断することは極めて困難な場合があります。このため個体差の影響が少ない唇弁(リップ)の構造、中央弁(Midlobe)およびカルスの形状が種の同定にしばしば用いられます。写真14は現在主な営利品種として販売されている胡蝶蘭の原種となったP. amabilisです。原種はすべて白色の花被片で、リップは黄色と白色ですが、青のストライプがリップ側弁入るものも稀に見られます。胡蝶蘭のなかでP. amabilisは台湾からタイ、マレーシヤ、、インドネシヤ、オーストラリアに至る最も広く分布する種です。鬱蒼とした密林のなかに生息するよりは比較的開けた山地林に着生し、花茎を1m以上伸ばして多くの花を付けます。園芸では夜間18C、昼間25C程度が続くと花茎を発生し、凡そ2か月で蕾から開花に至ります。 写真では台湾からボルネオ、Irjaに至る地域に分布する6つの花形状を示したものです。一見同じように感じられますが、地域によって花弁とリップに固有の形態が見られ、台湾およびインドネシア・スマトラ産では、花弁(中央の花びら)が楕円卵形で丸みあり、インドネシア・ジャワおよびイリジャでは花弁の付け根がやや細く突き出た菱形に似た形状に変化し、またボルネオ島では扇状となります。写真の映像から判断される花弁形状の地域差異は、台湾を北限として南下するに従って、丸みを帯びた形から花弁の付け根が細く伸びる菱形に代わり、また花弁サイズも大きくなる傾向が見られます。輪花数は台湾種は多く、南下するに従って少なくなります(台湾にはP. amabilisは生息しないという説があります)。 写真15は写真14のP. amabilisをその配列順にリップ部だけを切り取り、さらに側弁(Lateral lobe)を除いたものです。中央弁の基部にある左右の突起をもった小斑点をもつ黄色い塊がカルスです。 Phalaenopsis亜種のPhalaenopsis節ではカルスは1組となります。リップは同一種であるため、それぞれは類似していますが、カルスの形状、カルス側にある中央弁の、左右に手を広げたような3角形状の突起や、中央弁先端から伸びるヒゲなどの形状からも凡その地域が判断できます。例えば、カルスの突起部の形状、手の部分の有無(写真の種にはすべてありますがオーストラリア産には無いものがある)と位置、中央部の長さや面積など、確かにそれぞれの形状は微妙に違いがあります。(カルス形状と種の同定例は後節で詳細に取り上げます)。
以上のように花形態は種としての特徴は無論のこと、特に広域分布種は、1種のみであっても個体差や地域差の多様な情報を含んでおり、さらにそのいずれにも含まれないような交雑の可能性の謎解きなど、その収集は原種ならではの面白さがあります。また変種(form)を花柄だけでなく、匂い、葉、花茎などからも区別すれば、さらに多くの変種が生まれると思われます。 | ||||

|
葉形態の地域差 葉の色や形状は地域によって異なることがあります。写真16に2例を示します。
実生種に比べて肥料や水分補給の点でより不利な状況にある野生種がより大きな葉サイズとなる原因として筆者は乾季にあると推測しています。着生植物にとり可能な限り多くの水分の葉内への貯蔵が生存する上で重要でありその結果ではないかというものです。人工栽培では水と肥料は定期的に与えられており、大きくなる必然性がなく、それよりは早く開花し子孫を残すことが優先と考えています。自然界と人工栽培の違いはaphyllae亜属にも見られ、人工栽培では十分な水分補給が与えられるため落葉しません。これも環境に適応した生態と思われます。 | ||||

|
判定が困難な種 1.P. cornu-cervi Group胡蝶蘭には花柄や形態が極めて類似し、同定が困難な種(主にPolychilosやAphyllae亜種)があります。例えば下記のPolychilos亜属に含まれるP. borneensis、P. cornu-cervi、P. pantherina、 P. lamelligeraなどのそれぞれの種は花柄や花被片形状からでは判定は困難です。またこれらの一部を同一種と見なす考えもあります。 写真17は上記の種それぞれの花被片を示したもので、写真映像の花柄や形状を粒さに見れば、それぞれに個性を感じますが、何れの種もリップ中央弁形状を除き、相互に個体差の範囲で類似したものがあり明確な区別は困難です。  Fig. 17 Polychilos亜属 写真それぞれの種別判定するにあたり、写真の配列にそれぞれ対応させたリップおよびカルス形状を写真18および19に示します。写真19ではリップの左右の側弁をカルス形状が見やすいように取り外しています。 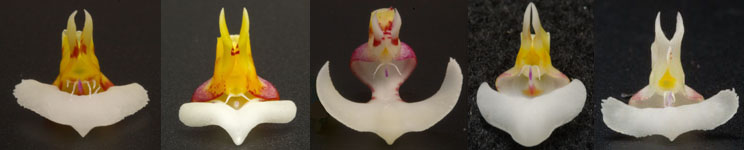 Fig.18 中央弁先端形状(Fig.17に対応)  Fig. 19. カルス構造(Fig.17に対応) 写真18および19の特徴を映像の視点から表6に整理します。表6の各項に関し、「中央弁」は写真の白い突き出した部分(1)を、「中央弁Pad」は中央弁の中心部(先端寄り)表面(2)、「中央弁長さ」は先端から基部までの長さ(3)、「棒状突起」は中央弁基部から突き出た先端が紫色の突起部(4)、「カルス形状」はリップ基部に最も近い(posterior)カルス(5)をそれぞれ指します。
映像から分析すると、明らかにP. lamelligeraとP. pantherinaは他の種と、中央弁(1)とカルス(central)形状に大きな特徴があります。カルスからは写真左のP. cornu-cerviの腺状突起(多数の小さな腺の集まり)が他種(2分岐突起)とは異なります。写真右の不明種はインドネシアからの入荷ですが、恐らくボルネオ島カリマンタン産と思われます。これまでの分類にはない特徴があります。それらはP. pantherinaと同じように中央弁は左右端を上部に反らした翼型となっているにも関らず、中央弁の長さ(3)は短くP. cornu-cervi同様にリップ基部に密着しています。またこの種の中央弁のPad上(2)はP. pantherinaと同形である反面、その凸部の上にはP. pantherinaに見られる僅かな細毛(写真では見ずらいため個別種説明のサイトを参照)がありません。P. cornu-cerviとP. pantherinaとの交雑種の可能性があるような形状といえます。それではその匂いはどこから来るのかという問題があります。 一方、これら種に関する分類学的情報はP. pantherinaを除いて極めて曖昧であり、著名なPhalaeniopsisのWebサイトにおいてもP.lamelligeraとP.borneensisに関しては花の映像が定まっていません。すなわち、写真19においてリップ形状がP.lamelligeraのもの、あるいはunknownの特徴をP.borneensisとしている等です。E.A. Christenson, "PHALAENOPSIS, A MONOGRAPH"においても、園芸種としてはP. cornu-cerviは広域に分布し一般的な種であるにもかかわらず、リップ解説図が取り上げられていません。これはP. cornu-cerviが地域差や個体差等(実生ではそれらの交雑を含め)を生んで多様化し、未だ学術分類的に整理されていないのではないかと思われます。前記著書にはP. lamelligeraはP. cornu-cerviと異なる種と見なす根拠がないとされています。しかし映像からは中央弁およびカルス形状が大きく異なり、写真からは同一種とは思えません。また香りを持つ点でも異なります。もし写真と名称が誤りであるのであれば、それでは写真のP.lamelligeraは新しい種なのかという問題が生じてしまいます。今後の研究が期待されます。 表7に分類や交雑関係の解明が難しい種を記載します。表以外ではP. amabilisやP. equestrisも含まれるかも知れません。
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||